Non sarebbe necessario scrivere una nota all’articolo pubblicato dall’Osservatorio Malattie Rare (O.ma.r.) sulla malattia di Lyme alcuni giorni fa [1], in cui si liquidano i possibili esiti cronici della patologia in parola come non esistenti o riconducibili ad altre patologie, “non ultime le patologie psichiatriche”. Non sarebbe necessario, ho scritto, perché coloro che si occupano professionalmente della malattia di Lyme, nonché i pazienti, sono consapevoli che in realtà le possibili sequele della infezione acuta da Borrelia burgdorferi sono ben documentate in letteratura e costituiscono un problema di enorme portata su cui si concentra oggi molta ricerca, spesso di alto profilo [2].
E allora perché questa preterizione? Perché un articolo divulgativo pubblicato da O.ma.r. ha notevole risonanza e – se incompleto o impreciso – rischia di alimentare false credenze tra i neofiti. L’articolo in questione è encomiabile nel divulgare nozioni preziose sulla fase acuta della malattia e nel mettere in guardia contro test e trattamenti non provati, ma è del tutto fuorviante nella parte dedicata alle sequele croniche (penultimo paragrafo).
La malattia di Lyme risponde alle attuali cure antibiotiche nel 90% dei casi. Questo significa che un decimo di coloro che ogni anno, in estate, contrae la malattia per il morso di un vettore (principalmente l’Ixodes ricinus in Europa) andrà incontro a una condizione cronica (cioè con durata superiore ai sei mesi) e debilitante, nota in seno alla comunità scientifica con il nome di post-treatment Lyme disease syndrome, PTLDS [3], [4]. Il nome scelto (che si potrebbe tradurre come Sindrome della malattia di Lyme dopo trattamento) sta a indicare che i pazienti sperimenteranno sintomi, nonostante i trattamenti della fase acuta.
La causa della PTLDS è al momento non nota. Alcuni studi supportano l’ipotesi che ci sia una disfunzione immunitaria in questi pazienti. Due studi hanno dimostrato la presenza di anticorpi contro il sistema nervoso centrale nella metà dei pazienti PTLDS [5], [6], solo per citare i più recenti. Per approfondimenti su questo argomento si legga qui e qui. La ricerca in questo campo continua [7].
Altri gruppi hanno dimostrato la persistenza del patogeno – dopo trattamento – sia nel modello animale della malattia di Lyme [8], [9], [10], che negli esseri umani [11], [12]. Presso la Columbia University è stata avviata una raccolta di tessuti provenienti da individui deceduti, che abbiano avuto una ben documentata infezione da Borrelia burgdorferi (link) proprio per indagare ulteriormente questo aspetto.
Numerosi sforzi e investimenti sono stati profusi recentemente nella ricerca di nuovi agenti antimicrobici per questa infezione, nella speranza di scongiurare le sequele croniche, da parte della Università di Stanford [13], [14], della Università Johns Hopkins [15], [16], [17], della Università Northeastern [18].
Nell’articolo di O.ma.r si legge che la sindrome in questione – chiamata impropriamente post-Lyme dall’autore – è caratterizzata da “sintomi soggettivi – e dunque non quantificabili – quali affaticabilità e difficoltà a concentrarsi”. A questo proposito è quasi superfluo ricordare che molti sintomi sono soggettivi, finché l’ingegneria non ci offre uno specifico strumento di misura: si pensi alla risonanza magnetica nella sclerosi multipla o all’elettroencefalogramma nella epilessia. In secondo luogo, questi sintomi sono solo in parte soggettivi, infatti i pazienti PTLDS presentano alterazioni quantificabili nel sistema immunitario [5], [6], nella espressione genica [19], nel metabolismo [20], etc.
Per quanto riguarda il riferimento all’ambito psichiatrico, vale la pena fare delle considerazioni. E’ senz’altro vero che le principali patologie psichiatriche (i disturbi dell’umore da un lato e le psicosi dall’altro) contemplano gli episodi infettivi come fattore di rischio [21], [22], tuttavia la PTLDS semplicemente non si sovrappone alle patologie psichiatriche, né nella presentazione clinica, né nella epidemiologia, per questo è una categoria nosografica a sé stante (vedi qui). Per fare un esempio concreto: negare l’evidenza, come sembra fare questo articolo, è una delle possibili manifestazioni della schizofrenia paranoide [23], ma né questo tipo di sintomi né altri sintomi patognomonici per malattie psichiatriche sono menzionati nella definizione di caso della PTLDS [4].
Paolo Maccallini
Riferimenti
Orzes, E. (2019, Nov. 7). Malattia di Lyme, attenzione alle false credenze. O.ma.r. (link)
Instutute of Medicine (2011, Apr. 20). Critical Needs and Gaps in Understanding Prevention, Amelioration, and Resolution of Lyme and Other Tick-Borne Diseases: The Short-Term and Long-Term Outcomes – Workshop Report. Cap. 7 (link).
Centers for Disease Control and Prevention. Post-Treatment Lyme Disease Syndrome. (link).
Aucott, J., Crowder, L., & Kortte, K. (2013). Development of a foundation for a case definition of post-treatment Lyme disease syndrome. Int J Infect Dis, 17(6), p. e443-e449. (link).
Chandra A, Wormser GP, Klempner MS, et al. Anti-neural antibody reactivity in patients with a history of Lyme borreliosis and persistent symptoms. Brain Behav Immun 2010;24:1018–24 (link).
Jacek E, Fallon BA, Chandra A, Crow MK, Wormser GP, Alaedini A. Increased IFNalpha activity and differential antibody response in patients with a history of Lyme disease and persistent cognitive deficits. J Neuroimmunol 2013;255:85–91 (link).
Maccallini, P; Bonin, S; Trevisan, G. Autoimmunity against a glycolytic enzyme as a possible cause for persistent symptoms in Lyme disease. Med Hypotheses. 2018 Jan. (link).
Hodzic E, Feng S, Holden K, Freet KJ, Barthold SW. Persistence of Borrelia burgdorferifollowingantibiotictreatment in mice. Antimicrob Agents Chemother. 2008 May; 52(5) (link).
Yrjänäinen H, Hytönen J, Hartiala P, Oksi J, Viljanen MK. Persistence of borrelial DNA in the joints of Borreliaburgdorferi-infected mice after ceftriaxone treatment. APMIS. 2010 Sep 1;118(9) (link).
Embers, M et al. Persistence of Borrelia burgdorferi in Rhesus Macaques following Antibiotic Treatment of Disseminated Infection. PLoS One. 2012; 7(1). (link).
Marques, A. et al. Xenodiagnosis to detect Borreliaburgdorferi infection: a first-in-human study. Clin Infect Dis. 2014 Apr; 58(7):937-45 (link).
Sapi, E. et al.The Long-Term Persistence of Borrelia burgdorferi Antigens and DNA in the Tissues of a Patient with Lyme Disease. Antibiotics 2019, 8(4), 183. (link).
Wagh D, Pothineni VR, Inayathullah M, Liu S, Kim KM, Rajadas J. Borreliacidal activity of Borrelia metal transporter A (BmtA) binding small molecules by manganese transport inhibition. Drug Des Devel Ther. 2015 Feb 11;9:805-16. (link).
Venkata Raveendra Pothineni et al. Identification of new drug candidates against Borrelia burgdorferi using high-throughput screening.Drug Des Devel Ther. 2016; 10: 1307–1322. (link).
Feng J, Wang T, Shi W, Zhang S, Sullivan D, Auwaerter PG, Zhang Y. Identification of novel activity against Borreliaburgdorferi persisters using an FDA approved drug library. Emerg Microbes Infect. 2014 Jul; 3 (7). (link).
Jie Feng, Megan Weitner, Wanliang Shi, Shuo Zhang, David Sullivan, and Ying Zhang. Identification of Additional Anti-Persister Activity against Borrelia burgdorferi from an FDA Drug Library. Antibiotics (Basel). 2015 Sep; 4(3): 397–410. (link).
Feng J, Auwaerter PG, Zhang Y. Drug combinations against Borreliaburgdorferi persisters in vitro: eradication achieved by using daptomycin, cefoperazone and doxycycline. PLoS One. 2015 Mar 25;10(3). (link).
Sharma B, Brown AV, Matluck NE, Hu LT, Lewis K. Borreliaburgdorferi, the Causative Agent of Lyme Disease, Forms Drug-Tolerant Persister Cells. Antimicrob Agents Chemother. 2015 Aug;59(8):4616-24. (link).
Jerome Bouquet et al. Longitudinal Transcriptome Analysis Reveals a Sustained Differential Gene Expression Signature in Patients Treated for Acute Lyme Disease. mBio. 2016 Jan-Feb; 7(1). (link).
Fallon BA et al. Regional cerebral blood flow and metabolic rate in persistent Lyme encephalopathy. Arch Gen Psychiatry. 2009 May;66(5):554-63. (link).
Benros ME et al. Autoimmune diseases and severe infections as risk factors for mood disorders: a nationwide study. JAMA Psychiatry. 2013 Aug;70(8): 812-20. (link).
Benros ME et al. Autoimmune diseases and severe infections as risk factors for schizophrenia: a 30-year population-based register study. Am J Psychiatry. 2011 Dec;168(12). (link).
Rajiv Tandon et al. Definition and description of schizophrenia in the DSM-5.Schizophrenia Research, Volume 150, Issue 1, October 2013, Pages 3-10. (link).
Queste sono solo alcune note raccolte dal discorso che Mark Davis ha pronunciato in occasione del Community Symposium tenutosi nell’agosto scorso (2017) a Stanford (video). Nei paragrafi 2, 3, 4 e 5 introdurrò alcune nozioni di base sui recettori delle cellule T (T cell receptors: TCR); il paragrafo 6, attraverso riferimenti al video già menzionato e a tre articoli pubblicati da Davis et al. nel corso degli ultimi quattro anni, descrive una nuova tecnologica sviluppata da Mark Davis e colleghi. Questi cenni preliminari dovrebbero auspicabilmente fornire i mezzi per comprendere a pieno la portata dei dati pilota presentati da Mark Davis a proposito dell’attività delle cellule T nella ME/CFS (paragrafo 7) e nella malattia di Lyme cronica (paragrafo 8), mostrando perché tale tecnologia prometta di divenire una sorta di test universale per qualsiasi tipo di infezione o patologia autoimmune, nota o sconosciuta.
2. Cellule T
I linfociti T sono una tipologia di leucociti (o globuli bianchi), vale a dire la componente cellulare del nostro sistema immunitario. La gran parte dei linfociti T in circolo è rappresentata da linfociti T helper (T helper cells:Th cells) e da linfociti T citotossici (cytotoxic T lymphocytes: CTL). Mentre la funzione dei linfociti T helper è quella di regolare l’attività degli altri leucociti attraverso la produzione di un’ampia gamma di trasmettitori chimici (le citochine, cytokines), le CTL sono coinvolte direttamente nella soppressione delle cellule ospiti infette. I linfociti T appartengono al ramo cosiddetto adattivo del sistema immunitario, assieme alle cellule B (le fabbriche di anticorpi), e, in quanto tali, il loro compito è quello di garantire una difesa specifica, su misura, contro gli agenti patogeni: per contrastare uno specifico agente patogeno, il nostro sistema immunitario può schierare in campo non solo anticorpi specifici ma anche specifici linfociti T (Th cells e CTL). Il ramo innato del sistema immunitario, invece, (nel quale rientrano le cellule natural killer, i macrofagi, le cellule dendritiche, i mastociti…) è in grado di fornire soltanto una difesa aspecifica, una prima linea di risposta immunitaria.
3. Recettori dei linfociti T
I linfociti T sono in grado di andare alla ricerca di specifici patogeni grazie a una molecola espressa sopra la propria superficie, chiamata recettore del linfocita T (TCR). Nella figura 1 si può vedere una schematica rappresentazione del TCR e del meccanismo in virtù del quale il linfocita T riconosce il proprio target. Gli antigenti (proteine) degli agenti patogeni vengono indicati ai linfociti T da altre cellule del nostro corpo: vengono esposte sopra molecole chiamate Complesso Maggiore di Istocompatibilità (MHC), che si trova espresso sulla membrana esterna. Se un dato antigene mostra compatibilità con il TCR di uno specifico linfocita T, tale linfocita T si attiva e comincia a proliferare (espansione clonale, clonal expansion). Le due catene principali (α and β) sono assemblate combinando la trascrizione di segmenti di gene, ognuno dei quali ha copie multiple, leggermente diverse fra loro: in altre parole, i TCR vengono assemblati a partire da peptidi scelti a caso da un insieme di diverse alternative possibili. Questo comporta un repertorio di 10^15 diversi possibili TCR (Mason DA 1998). Ogni linfocita T mostra un solo tipo di TCR.
4. Cellule T helper
Le cellule T helper sono programmate per riconoscere esclusivamente antigeni esposti dalle molecole MHC di seconda classe (II): questa classe di MHC viene espressa sulla membrana esterna di alcuni leucociti, principalmente le cellule dendritiche, le cellule B e i macrofagi (tutte assieme dette “cellule che presentano l’antigene”, antigen presenting cells: APC). Le molecole MHC II legano il TCR delle cellule T helper grazie al peptide CD4 (espresso unicamente dalle cellule T helper). L’antigene presentato dalle molecole MHC è un peptide lungo 13-17 amminoacidi (Rudensky, et al., 1991) (figura 2).
5. Linfociti T citotossici
I TCR espressi dai linfociti T citotossici (CTL) possono legare solo antigeni esposti dalle molecole MHC di prima classe (I), che si trovano nella membrana esterna di qualunque cellula del nostro corpo. La glicoproteina CD8 è la molecola che rende i TCR espressi dalle CTL specifici per il MHC I. Mentre gli antigeni esposti dalle APC appartengono a patogeni raccolti sul campo di battaglia di passate infezioni, i peptidi esposti dal MHC I di una specifica cellula appartengono a patogeni che hanno fatto ingresso nella cellula stessa, e pertanto costituiscono la prova di un’infezione intracellulare ancora in atto (figura 3). Nel momento in cui un CTL riconosce un antigene che combacia con il proprio TCR, il CTL iduce l’apoptosi (morte programmata) della cellula che mostra l’antigene. Gli antigeni esposti dal MHC I sono peptidi che vanno dagli 8 ai 10 amminoacidi (Stern, et al., 1994).
Figure 3. Una cellula infetta espone un antigene virale sul proprio MHC I. Il TCR di un CTL si lega a questo peptide ed invia un segnale interno diretto al suo proprio nucleo, il quale risponde attivando l’apoptosi (attraverso il rilascio di granzimi, ad esempio) della cellula infetta (disegno di Paolo Maccallini).6. Il test immunologico universale
Nel corso del suo discorso, Mark Davis illustra alcuni concetti base sul sistema immunitario, prima di passare a introdurre i nuovi, entusiasmanti dati riguardo alla ME/CFS e alla Lyme cronica (o post-treatment Lyme disease syndrome: PTLDS). Contestualmente, però, dedica alcuni minuti alla descrizione di un complesso nuovo test che teoricamente renderebbe possibile estrapolare tutte le informazioni contenute nel repertorio di TCR presenti – in un dato momento – nel sangue di un essere umano. Un test del genere – che chiamerei “test immunologico universale” – avrebbe la capacità di determinare se un paziente presenta un’infezione in corso (e, nel caso, indicare il patogeno coinvolto) oppure una malattia autoimmune (anche qui specificando la natura dell’autoantigene, ossia il tessuto attaccato dal sistema immunitario). A quanto mi è dato di comprendere, il test richiede tre passaggi, che elenco nelle sezioni seguenti.
6.1. Primo step: sequenziamento del TCR
Come già spiegato nel paragrafo 3, quando un linfocita T incontra un peptide a cui è compatibile, comincia a proliferare; pertanto, nel sangue di un paziente con infezione in corso (o con reazione contro il proprio organismo, cioè con reazione autoimmune) è possibile trovare molteplici copie di linfociti T che esprimono il medesimo TCR: a differenza dei controlli sani, nei quali circa il 10% delle CD8 totali è rappresentato da copie di pochi diverse linfociti T (figura 4, prima linea), nei pazienti affetti da Lyme incipiente – un esempio di infezione attiva – o sclerosi multipla (MS) – un esempio di malattia autoimmune – abbiamo una massiccia clonazione di alcune linee di CTL (figura 5, seconda e terza riga, rispettivamente). Il primo step del test immunologico universale starà allora nell’identificazione dell’esatta sequenza di TCR espressa dai linfociti T presenti nel sangue, come si legge in Han A et al. 2014, dove troviamo descritto il sistema per sequenziare i geni delle catene α e β di un dato linfocita T. Tale approccio permette di costruire grafici come quello in figura 4 e quindi permette di determinare se il paziente presenti in atto un’attività anomala dei linfociti T oppure no. Qualora si riscontri un fenomeno di espansione clonale, è legittimo ipotizzare che stia avendo luogo o un’infezione o una condizione autoimmune di qualche sorta.
Figure 4. Ogni cerchio rappresenta un paziente. Nella prima riga vediamo quattro controlli sani, che non presentano affatto espansione clonale delle cellule CD8 (come nel primo paziente da sinistra) oppure la presentano in maniera assai moderata (come indicato dalle porzioni in blu, bianco e grigio). Nella seconda riga troviamo invece quattro pazienti con malattia di Lyme attiva (fase incipiente) e possiamo ben notare come ciascuno di loro, nessuno escluso, presenti espansione clonale di solo tre diverse T cells (porzioni in rosso, blu e arancione). Nella terza riga, infine, abbiamo quattro pazienti affetti da MS, le cui cellule CD8 sono per maggior parte rappresentate da cloni di una selezione ristretta di T cells. Fonte: slide proposte da Mark Davis durante il Community Symposium.
6.2. Secondo step: raggruppamento dei TCR
Mark Davis e colleghi hanno realizzato un software capace di identificare i TCR che condividono il medesimo antigene, sia in un singolo individuo che trasversalmente a un gruppo. L’algoritmo è stato denominato GLIPH (grouping of lymphocyte interaction by paratope hotspots) ed ha dato prova di poter raggruppare – per fare un esempio – i recettori dei linfociti T CD4 di 22 soggetti con infezione da M. tuberculosis latente in 16 gruppi distinti, ognuno dei quali comprende TCR provenienti da almeno tre individui (Glanville J et al. 2017). Cinque di questi gruppi sono riportati nella figura 5. L’idea sottostante è che TCR che appartengono allo stesso raggruppamento reagiscano allo stesso complesso peptide-MHC (pMHC).
Figure 5. Cinque gruppi di TCR provenienti da 22 diversi pazienti affetti da turbercolosi latente, raggruppati grazie al GLIPH. La prima colonna da sinistra riporta l’identificativo dei TCR; la seconda l’identificativo dei pazienti. Le CDR per le catene β e α si trovano, rispettivamente, sulla terza e sulla quinta colonna. Tratto da Glanville J et al. 2017.
6.3. Terzo step: ricerca degli epitopi
Come abbiamo visto, questa nuova tecnologia consente di rilevare se sia in atto un’espansione clonale di linfociti T sequenziando i TCR dal sangue periferico. Consente inoltre di raggruppare i TCR presenti in un singolo paziente o condivisi da più pazienti. Il passaggio successivo è quello di identificare a quale/i tipo/i di antigene ognuno di questi raggruppamenti reagisca. Infatti, se potessimo identificare degli antigeni comuni in un gruppo di pazienti dai sintomi comparabili nei quali si riscontri un’espansione clonale in atto e simili TCR, saremmo messi in grado di comprendere se il loro sistema immunitario stia attaccando un patogeno (e di identificare il patogeno) o se stia piuttosto attaccando un tessuto ospite e, qualora fosse questo il caso, di identificare il tessuto. Come già detto, il numero di possibili combinazioni per il materiale genetico dei TCR è calcolato attorno ai 10^15, ma il numero dei possibili epitopi di cellule Th è circa 20^15, che corrisponde a più di 10^19. Ciò implica che i TCR debbano essere in una qualche misura cross-reattivi se vogliono essere in grado di riconoscere tutti i possibili peptidi esposti dai MHC (Mason DA 1998). Il grado di tale cross-reattività e il meccanismo attraverso il quale viene ottenuta sono stati spiegati con esattezza da Mark Davis e colleghi in un recente articolo (Birnbaum ME et al. 2014), che ci fornisce il terzo step del test immunologico universale. Lo scopo di questa fase consiste nel prendere un dato TCR e trovare il repertorio dei suoi specifici antigeni (giova ripetere che, appunto, ogni TCR reagisce a più antigeni). Per comprendere come ciò sia possibile, guardiamo a uno degli esperimenti descritti nell’articolo più sopra citato. I ricercatori si sono concentrati su due TCR ben precisi (chiamati Ob.1A12 e Ob.2F3), clonati da un paziente con MS e noti per essere capaci di riconoscere i pepetidi 85-99 (figura 6) della proteina basica della mielina (MBP) esposti dall’ HLA-DR15. Hanno poi preparato un insieme di cellule di lievito che esprimono molecole HLA-DR15, ognuna caratterizzata da un diverso peptide formato da 14 amminoacidi, con amminoacidi fissi esclusivamente alle posizioni 1 e 4, dove il peptide è ancorato al MHC (figura 6, sinistra). Quando alla coltura di cellule di lievito che esprimono complessi pMHC vengono aggiunte copie di Ob.1A12, queste legano solo con alcune di quelle e, come è possibile vedere dalla parte destra della figura 6, per ciascuna posizione degli epitopi legati dal Ob.1A12 esiste un amminoacido con maggior tasso di probabilità: ad esempio, l’epitopo Ob.1A12 tipico presenta preferibilmente alanina (A) in posizione -4, istidina (H) in posizione -3, arginina (R) in posizione -2, e così via. Da notare che istidina (H) in posizione 2 e fenilanina (F) in posizione 3 sono amminoacidi obbligatori per un epitopo di Ob.1A12.
Figure 6. Sulla sinistra: il peptide 85-99 della proteina basica della mielina (myelin basic protein, MPB) è risaputo essere un epitopo per il TCR Ob.1A12. In posizione 1 e 4 possiede due residui che gli consentono di legare con la molecola MHC. In posizione -2, -1, 2, 3 3 5 troviamo invece i residui che legano con il TCR. La seconda riga rappresenta l’epitopo generico della libreria peptidica utilizzata per identificare il grado di cross-reattività tra tutti i possibili epitopi di Ob.1A12.A destra: la probabilità di ciascun amminoacido per ciascuna posizione è rappresentata da sfumature di viola.Come potete vedere, l’istidina (H) in posizione 2 e la fenilalanina (F) in posizione 3 sono amminoacidi obbligatori affinché un epitopo sia reattivo con Ob.1A12.Da (Birnbaum ME et al 2014).
La tabella sulla destra della figura 6 rappresenta, infatti, una tabella di sostituzione (substitution matrix) di dimensioni 14×20, uno strumento impiegato per scansionare il database dei peptidi in modo da trovare, tra tutti i peptidi conosciuti espressi da creature viventi, tutti i possibili epitopi specifici per Ob.1A12. Le matrici di sostituzione vengono solitamente utilizzate nel cosiddetto allineamento di peptidi (peptide alignment), una tecnica che punta all’identificazione di similitudini tra peptidi. Tali matrici sono basate su considerazioni di tipo evoluzionistico (Dayhoff, et al., 1978) o sullo studio delle regioni conservate delle proteine (Henikoff, et al., 1992). Ma la ricerca degli epitopi specifici di un dato TCR richiede (come abbiamo visto per Ob.1A12) una matrice di sostituzione costruita ad hoc per quel TCR: ogni TCR richiede la propria matrice di sostituzione, ottenuta incubando cellule T esprimenti quel TCR con una coltura di lieviti che espongono sui propri MHC una grande varietà di peptidi casuali, e analizzando poi i dati ricavati dall’esperimento. Quindi, un processo piuttosto complesso! Nel caso di Ob.1A12, questo processo ha portato a 2330 peptidi (incluso MBP), mentre la matrice di sostituzione specifica per Ob.2F3 ha trovato 4824 epitopi all’interno dell’intero database di peptidi. Questi peptidi includevano sia proteine non umane (batteriche, virali…) che peptidi umani. Per 33 di loro (26 non umani e 7 proteine umane), questo gruppo di ricercatori ha eseguito un test per verificare direttamente la previsione: 25/26 dei peptidi ambientali e 6/7 dei peptidi umani hanno indotto la proliferazione di cellule T che esprimono il TCR Ob.1A12 e/o il Ob.2F3, e questa è una prova della validità di questa analisi! Questi 33 peptidi sono riportati nella figura 7. Questo è l’ultimo passaggio del test immunitario universale, quello che dal TCR conduce agli epitopi. Come avete visto, un enorme insieme di diversi peptidi da diverse fonti reagisce con un singolo tipo di TCR; in altre parole, la cross-reattività è una proprietà intrinseca del TCR. Ciò significa anche che i test di trasformazione linfocitaria (LTT), ampiamente utilizzati in Europa per l’individuazione di infezioni da Borrelia burgdorferi e altri patogeni, comportano un rischio elevato di risultati falsi positivi e richiedono un processo di validazione sperimentale e teorica, attualmente mancante.
Figura 7. Una serie di 33 peptidi che si suppongono essere epitopi specifici sia per Ob.1A12 che per Ob.2F3. Tratto da Birnbaum ME et al. 2014.
Siamo ora pronti ad apprezzare appieno i dati pilota che Mark Davis ha presentato al Symposium sull’espansione clonale delle cellule T CD8 nella ME/CFS e nella Lyme cronica.
7. “We have a hit!”
Mark Davis, insieme a Jacob Glanville e José Montoya, hanno sequenziato TCR dal sangue periferico di 50 pazienti ME/CFS e 49 controlli (primo passo del test immunitario universale, ricordate?), quindi li hanno raggruppati usando l’algoritmo GLIPH (secondo passo).Hanno trovato 28 cluster, ciascuno costituito da più di 2500 sequenze simili, e ogni cluster raccoglie sequenze multiple dallo stesso individuo e sequenze (che sono forse più importanti) da pazienti diversi (figura 8). Il cluster che ho cerchiato in rosso, ad esempio, è una raccolta di oltre 3000 TCR simili. La presenza di questi ampi cluster nei pazienti ME/CFS, rispetto ai controlli sani, rappresenta una prova indiretta di una risposta specifica delle cellule T a un trigger comune in questo gruppo di pazienti, che potrebbe essere un agente patogeno o un tessuto del corpo (o tutti e due).
Figura 8. Nella ME/CFS le sequenze di TCR ricavati da 50 pazienti formano 28 raggruppamenti che presentano più di 2500 sequenze simili – cosa che assolutamente non avviene nei controlli sani. Questo fa pensare ad una qualche risposta immunitaria ad un patogeno o ad un tessuto umano (o entrambi). Fonte: slide proposta da Mark Davis durante il Community Symposium.
Tra questi 50 pazienti ME/CFS, Davis e colleghi hanno selezionato 6 pazienti con geni HLA simili (figura 9, sinistra), 5 femmine tra loro, e hanno studiato i loro TCR più in profondità. Nella metà destra della figura 9, è possibile vedere per ciascun paziente il grado di espansione clonale delle CTL. Ricordate che nei controlli sani solo circa il 10% dei CTL è composto da cloni di alcune cellule (figura 4, prima riga), mentre qui vediamo che circa il 50% di tutti i CTL è composto da cloni. Quindi, una “marcata espansione clonale” delle cellule T CD8, come ha detto Davis.
Figura 9. A sinistra: sono stati selezionati 6 pazienti ME/CFS con HLA simili. Sulla prima colonna da sinistra sono stati riportati gli identificativi dei pazienti; la seconda colonna ci informa sull’età di ciascuno; la terza sul genere; la quarta avvisa di eventuali esposizione a citomegalovirus; la quinta riguarda i geni del MHC I. A destra: l’analisi dell’espansione clonale delle cellule T CD8 per ognuno dei pazienti. L’espansione clonale è marcata (circa al 50%), se comparata a quella dei controlli sani (circa al 10%).
Le sequenze delle catene α e β dei TCR di tre dei sei pazienti (pazienti L4-02, L4-10 e L3-20) sono riportate in figura 10 (ho verificato che effettivamente si tratta di catene α e β di TCR umani, inserendole in BLAST).
Figura 10. Catene β (prima colonna) e rispettive catene α (quinta colonna) provenienti da tre pazienti ME/CFSchains (L4-02, L4-10, and L3-20, ultima colonna).
Quindi, abbiamo visto finora i primi due passaggi del test immunitario universale. E il terzo passo? Nel suo discorso, Mark Davis non ha presentato alcun particolare epitopo, ha solo mostrato una diapositiva con quella che probabilmente è la selezione degli epitopi dalla libreria discussa nel paragrafo 6.3 da parte di uno dei TCR riportati in figura 10. Questa selezione è riportato in figura 11, ma da quella foto non è possibile raccogliere alcuna informazione sull’identità di questi epitopi. Come probabilmente ricorderete dal paragrafo 6.3, l’analisi dei peptidi selezionati da un TCR nella libreria di peptidi consente l’identificazione di una matrice di sostituzione che può essere utilizzata per selezionare tutti i possibili epitopi di quel TCR specifico, dal database delle proteine. Quest’ultima fase cruciale deve essere ancora eseguita, o è già stata eseguita, ma Davis non ha comunicato i risultati preliminari durante il suo discorso. Recentemente sono state messe a disposizione nuove risorse dalla Open Medicine Foundation, affinché questa ricerca promettente possa essere ulteriormente perseguita (R). Lo scopo qui, come già detto, è di trovare l’antigene che innesca questa risposta delle cellule T. Come ha detto Mark Davis, potrebbe essere un antigene di un agente patogeno specifico (forse un patogeno comune che va e viene) che suscita una risposta immunitaria anomala che finisce per colpire alcuni tessuti ospiti (microglia, per esempio), portando così attivazione immunitaria che è stata recentemente segnalata da Mark Davis stesso e altri in ME/CFS (Montoya JG et al. 2017). L’idea di un patogeno comune che innesca una risposta immunitaria patologica non è nuova in medicina, e la febbre reumatica (RF) è un esempio di una tale malattia: la RF è una malattia autoimmune che attacca il cuore, il cervello e le articolazioni ed è generalmente innescata da uno streptococco che infetta la gola (Marijon E et al. 2012). L’altra possibilità è, naturalmente, quella di un’infezione in corso di qualche tipo, che deve ancora essere rilevata. Come detto (vedi par. 6.1), l’espansione clonale delle cellule T CD8 è presente sia nelle infezioni acute (come la malattia di Lyme) che nelle malattie autoimmuni (come la SM) (figura 4), quindi dobbiamo aspettare l’identificazione dell’antigene se vogliamo capire se l’attività del CTL è contro un agente patogeno e/o contro un tessuto del nostro corpo.
Figura 11. Nella figura possiamo osservare la selezione, che avviene in più momenti, di una serie di peptidi da parte di un particolare TCR proveniente da un paziente ME/CFS. La selezione ha luogo tra una enorme quantità di peptidi esposti dall’ HLA-A2 (MHC I) espresso da cellule di lievito. Ad ogni passaggio il numero di possibili peptidi si riduce.
8. La Lyme cronica esiste
È stato probabilmente trascurato il fatto che nel suo discorso, Mark Davis ha riportato anche dati molto interessanti sulla sindrome della malattia di Lyme post-trattamento (PTLDS, nota anche come malattia di Lyme cronica). In particolare, ha trovato un’espansione clonale marcata nelle cellule T CD8 di 4 pazienti PTLDS (circa il 40% dei CTL totali) come riportato nella figura 12: si consideri che in questo caso le fette blu rappresentano cellule T uniche, mentre tutte le altre fette rappresentano cloni! Tutto ciò che è stato detto sull’espansione clonale CD8 nella ME/CFS si applica anche in questo caso: potrebbe essere la prova di un’infezione in corso – forse la stessa B. burgdorferi, come suggerito da diversi modelli animali (Embers ME et al. 2017), (Embers ME et al. 2012), (Hodzic E et al. 2008), (Yrjänäinen H et al. 2010) – o una coinfezione (un virus?). Oppure potrebbe essere l’espressione di una reazione autoimmune innescata dalla infezione iniziale. Questo deve ancora essere scoperto, eseguendo il test immunitario universale completo, ma ciò che è già chiaro dalla figura 12 è che la PTLDS è una condizione reale, con qualcosa di veramente anomalo nella risposta immunitaria: la Lyme cronica esiste.
Figura 12. Espansione clonale di cellule T CD8 in quattro pazienti affetti da Lyme cronica. L’espansione clonale, che indica l’attività delle cellule T contro un patogeno o un tessuto ospite, è assai marcata.
9. Conclusioni
Mark Davis e altri ricercatori hanno sviluppato un test complesso che è in grado di sequenziare i TCR dai pazienti, raggrupparli in gruppi di TCR che reagiscono agli stessi antigeni e scoprire gli antigeni che hanno attivato quella particolare risposta delle cellule T. Questo test è una sorta di test immunitario universale che è teoricamente in grado di riconoscere se una persona (o un gruppo di pazienti) presenta una risposta immunitaria contro un agente patogeno o contro uno dei loro stessi tessuti (o entrambe le cose). Questo approccio ha già fornito dati pilota su una attivazione anomala delle cellule T CD8 nei pazienti ME/CFS e nei pazienti PTLDS e, si spera, identificherà il trigger di questa risposta immunitaria nel prossimo futuro. Se la ME/CFS è causata da un’infezione attiva, da una malattia autoimmune o da entrambe le cose, il test immunologico universale potrebbe essere in grado di dircelo. Questa nuova tecnologia è per l’immunologia, ciò che il sequenziamento dell’intero genoma è per la genetica, o la metabolomica è per le malattie molecolari: non cerca un particolare agente patogeno o una particolare malattia autoimmune. No, cerca tutte le possibili infezioni e disturbi immunitari, anche quelli che devono ancora essere scoperti.
A line of T cells (called Ob.2F3) expressing the same T cell receptor (TCR) from an MS patient was studied in 2014 and it was found to proliferate when incubated with 4824 different peptides. Thirty-three of them were further studied (see figure) and found to belong to both Homo sapiens and several different, unrelated microbes (Birnbaum ME et al. 2014). The taking home message here is that T cells are not specific to a single pathogen, they are highly cross-reactive, as it was already pointed out in this pivotal study: (Mason DA 1998). And this means that we can’t use lymphocyte transformation tests (LTTs) the way we do now.
I feel really frustrated when patients send me their LTTs and ask me to comment the results. I have to say that they have wasted their money and that these results are useless. I do hope that my blog can make a difference and stop this unfair commerce at the expenses of desperate folks.
Figure. A set of 33 peptides (both human and environmental) predicted to be specific epitopes for both Ob.1A12 and Ob.2F3. From (Birnbaum ME et al. 2014).
Quello che segue è il mio intervento durante il convegno nazionale sulla ME/CFS tenutosi a Thiene, tappa italiana dell’End ME/CFS Worldwide Tour. L’intervento è molto denso e veloce, ho dovuto condensare 4 anni di ricerche in 30 minuti. Qualcuno ha notato che sembravo dopato. Lo ero, letteralmente: ero alla fine di un lungo trattamento cortisonico e avevo assunto modafinil per la circostanza. Altrimenti non sarei riuscito. Le slide usate per questo intervento, insieme ad altre che non ho avuto modo di far vedere al convegno, sono disponibili qui. Sotto il video c’è un sommario del contenuto dell’intervento, e il momento del video in cui i vari argomenti sono stati trattati. Ringrazio Giuseppe Pozza per aver realizzato il video.
Note biografiche, criteri diagnostici e disturbi cognitivi (00:50)
Intolleranza ortostatica (08:39)
Citochine (09:45)
Citotossicità delle NK (11:15)
Disfunzioni metaboliche (12:40)
Anomalie del sistema nervoso centrale (20:50)
Anomalie del microbioma (22:40)
Analisi genetica (24:20)
La mia ricerca su Lyme e autoanticorpi (27:50)
Anticorpi anti-muscarinici e anti-beta adrenergici nella M/CFS (32:20)
Studi a cui ho partecipato come paziente, conclusioni e ringraziamenti (34:53)
Riascoltando il mio intervento ho provato stupore nel constare, forse per la prima volta, come la mia mente sia sopravvissuta. Solo io posso sapere cosa ho passato, nessuno sa che per gli ultimi 17 anni sono stato incapace di pensare per più del 90% del tempo. E non intendo incapace di risolvere sistemi di equazioni differenziali; no, intendo incapace di sostenere una conversazione o di leggere un libro.
Nonostante sia stata così colpita dalla malattia, nonostante sia stata privata di stimoli, nostante la solitudine estrema, i farmaci inutili e il consumarsi dei lustri, è sopravvissuta. Questo organo di un chilo e mezzo scarso che mi contiene tutto, che ha perso così tanto, che ho dato per spacciato tante volte, è sopravvissuto. La vita vuole vivere e non si arrende.
Per ottimizzare i test sierologici in termini di sensibilità e specificità, si adotta in Europa, quanto negli Stati Uniti, un algoritmo a due livelli (vedi Figura 1). Il primo livello è costituito da un test ad alta sensibilità (il test ELISA); coloro che sono negativi a questo test vengono considerati sieronegativi, coloro che sono positivi accedono a un test di secondo livello altamente specifico (il western blot), il cui scopo è quello di minimizzare il numero di falsi positivi (Wilske B. et al. 2007).
Figura 1. Algoritmo decisionale per il test a due livelli.
In Tabella 1 sono riportati i dati statistici relativi a un test a due livelli in cui il primo livello è rappresentato da un test ELISA con antigeni ottenuti da sonicato da cellula intera di Borrelia afzelii per le IgM, con aggiunta di VlsE ricombinate da B. afzelii, B. garinii e B. burgdorferi ss per le IgG; il secondo livello è invece costituito da un western blot i cui antigeni sono proteine estratte da B. burgdorferi ss (ceppo B31) e da Borrelia afzelii (ceppo PKo) con aggiunta di VlsE ricombinate da B burgdorferi ss (ceppo B31) (Branda JA. et al. 2013). Si può notare che con questa procedura si ha un 12% di falsi negativi nel gruppo neuroborreliosi e una percentuale di falsi positivi sempre minore del 2%.
Tabella 1. Sono riportati sensibilità, specificità, valore predittivo positivo (VPP) e valore predittivo negativo (VPN) per un western blot europeo. EM sta per eritema migrante, NB per neuroborreliosi, ACA per acrodermatite cronica atrofica. IgG e IgM sono considerate insieme. Da (Branda JA. et al. 2013) con modifiche.
A conclusione di questo paragrafo, si segnala che nel 2011 i CDC hanno modificato le loro raccomandazioni, includendo la possibilità dell’uso diretto di un immunoblot per le IgG, senza passare per il test di primo livello (R).
Donazione
Considera una donazione per sostenere questo blog.
Sul versante dei test, si parla di test di stimolazione linfocitaria, sia utilizzando le cellule T che utilizzando i macrofagi. Questi test sono in fase di sviluppo e quelli già venduti ai pazienti (in Germania ad esempio) non sono in realtà al momento utilizzabili.
Sul lato dei sintomi cronici: le cause ipotizzate sono persistenza della infezione, alterazioni immunitarie, modifiche nell’encefalo, coinfezioni non identificate (e quindi non trattate).
Fallon enfatizza il fatto che la persistenza della infezione non è da considerare la sola causa dei sintomi cronici. Da quello che sento ogni giorno, i pazienti sono dogmatici su questo punto: i sintomi cronici sono il prodotto di una infezione cronica. Può anche essere, in alcuni casi, ma per ora non si sa e tutti i tentativi di cura dei sintomi cronici sono risultati fallimentari: non ci sono farmaci approvati per questi pazienti. I medici spesso sono dogmatici, perché il dogma richiede il minimo sforzo cognitivo; i pazienti non dovrebbero fare lo stesso errore.
In generale io sono d’accordo con quello che dice Fallon, perché è molto onesto. Del resto lui non ha nulla da vendere e il suo scopo è perseguire la verità, avendo anche un congiunto malato.
Donazione
Considera una donazione per sostenere questo blog.
A new study from Columbia University, Stony Brook University and the CDC – that has seen the collaboration of Brian Fallon and W. Ian Lipkin, among others – has proposed a new serological test for Lyme disease and coinfections (Babesia microti, Anaplasma phagocytophilum, Ehrlichia chaffeensis, Rickettsia ricketsii and some viruses that are not present in Europe) (Tokarz R et al. 2018). Current serologic analyses consist of a two-tiered algorithm: a measure of serum activity against a main immunogenic protein of B. burgdorferi (whether it is VlsE or its peptide C6) is followed by a western blot, where the immune response to a set of several full-length proteins is performed. This method lacks sensitivity for early Lyme disease (Aguero-Rosenfeld ME et al 2005) and – according to studies on the animal model of Lyme disease – it might miss some cases of disseminated infection too (Embers ME, 2012), (Nicholas A, 2017), (Embers ME 2017). So, there is common agreement that a better test is urgently needed.
The quest for immunogenic peptides in Lyme patients: the set of proteins
Most B cells epitopes on non-denaturated proteins (i.e. proteins that conserve their tertiary structure) are believed to be conformational (Morris, 2007) but it is also true that in the average B cell epitope, a linear stretch of 5 amino acids is reported (Kringelum, et al., 2013). This means that it is conceivable to search for new immunogenic peptides in Lyme disease with the following method: each protein from Borrelia, known to be immunogenic in humans, is divided in peptides with a fixed length, then each of these peptides is exposed to sera from patients. Those peptides that strongly bind sera from patients are eligible as immunogenic peptides useful for diagnostic purposes. This is exactly what has been done in this study, and this elaborate analysis has been performed not only for Borrelia burgdorferi but also for the other tick-borne pathogens already mentioned. In particular, the length of the peptides has been fixed to be 12, and contiguous peptides have an overlapping of 11 amino acids. The set of immunogenic proteins chosen for each pathogen are reported in table 1.
Table 1. Set of proteins from various pathogen chosen by the Authors.
A new array of diagnostic peptides in Lyme disease
The analysis was conducted using sera from 66 Lyme patients (27 early Lyme, 19 with positive IgG Western blot, 10 with acute neuroborreliosis). The end result is the set of peptides reported in table 2. There is also a set of peptides specific for neuroborreliosis (table 3).
Table 2. Peptides on the second column of this table are specific immunogenic proteins in Lyme disease patients. In the first column proteins they belong to are collected.Table 3. Imunogenic peptides in neuroborreliosis.
Flagellin B: a new specific peptide for diagnostic purposes is born
It is widely recognized that reactivity of sera to Flagellin B (FlaB, p41) is common in healthy persons too (Chandra A et al. 2011). This is because this protein is highly conserved among bacteria, so there is cross-reactivity and FlaB is almost without utility for diagnostic purposes. That said, one of the results of this study is that peptide 211-223 fro FlaB is highly immunogenic in patients and is also not-cross-reactive with flagellar proteins from other bacteria (I have checked using this peptide as query sequence in a BLAST search among bacteria, with a word of 3 and with standard settings: no match has been found). So we have a new specific peptide for diagnostic purposes. In figure 1 this epitope is reported in yellow, on the 3D structure of Flagellin B.
Figure 1. Peptide 211-223 of flagellin B, in yellow. This structure is built by Modbase using the sequence P11089 and using flagellin of E. coli as a template.
The strange case of VlsE and C6
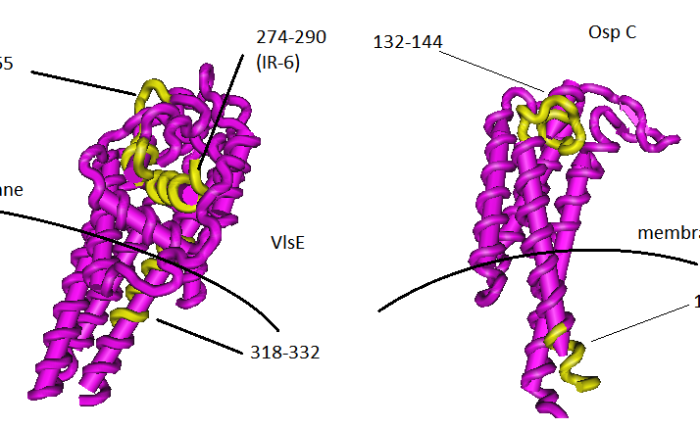
Vmp-like sequence expressed (VlsE) is a major immunogenic protein of Borrelia burgodorferi, widely used for the first-tier test of Lyme disease (Aguero-Rosenfeld ME et al 2005). This protein goes through a process of variation during mammalian infection that involves six variable regions (VR1-VR6) and it has been postulated that this gene recombination of the locus that codes for VlsE is responsible – at least in part – for the ability of B. burgodorferi of evading the immune system (Zhang JR et al. 1997). Nevertheless, six regions of this protein are invariable (IR1-6) and these ones have been studied as possible peptides to use for diagnosis. One of them, peptide IR6 (also called C6), has been used as a diagnostic tool (Liang FT et al. 1999). In the present study, the peptide that pops up from the analysis is the same C6 (peptide 274-290 in table 1), while in neuroborreliosis there are two more peptides on VlsE that appear to be useful markers. I have reported these three peptides on the 3D structure of VlsE, the one experimentally determined in (Eicken C et al. 2002), see figure 2. It is worth noting now that peptide 274-290 (C6) is buried inside the protein, so it is difficult to understand how it could be a B cell epitope since B cell epitope protrudes from the surface (Thornton, et al., 1986) and this is probably because B cell receptors need to bind their specific antigen, for B cell to be activated. This is the very first time that I realize that C6 is not surface exposed. So, how could the immune system generate an antibody to a hidden peptide? I have to think about that.
Figure 2. Immunogenic peptides on VlsE (left) and on OspC (right).
OspC and the buried epitope
Another weird case is that of OspC, where one of the two main epitopes found in patients with neuroborreliosis is buried in the cytoplasmic membrane of Borrelia burgdorferi (figure 2, right). This is an uncommon case, I guess, for the same reason mentioned above: BCRs need surface exposure to bind their specific epitope. The other peptide (132-144) appears to be a classic surface-exposed B cell epitope.
Quella che segue è una presentazione della Associazione Lyme Italia e Coinfezioni, costituitasi due anni fa e già distintasi in numerose attività a sostegno di coloro che sono affetti dalla malattia di Lyme e dei loro congiunti, e a favore della ricerca scientifica su questa patologia. Come molte altre persone, anche io ho accumulato un notevole debito di gratitudine nei confronti del Consiglio Direttivo dell’Associazione e della associazione tutta, e a loro esprimo il mio grazie e l’augurio di proseguire con la stessa tenacia e intraprendenza, ad meliora et maiora. (PM)
Nel dicembre del 2015 è nata, come prima realtà sul territorio nazionale per la specifica tematica, l’Associazione Lyme Italia e Coinfezioni grazie alla volontà di un gruppo eterogeneo di persone entrate in contatto con la Malattia di Lyme, o perché contratta direttamente, o perché familiari di malati, oppure ancora perché professionisti sanitari, come medici e veterinari.
Il bisogno di conoscere meglio la Malattia di Lyme e i luoghi dove curarsi, ha portato i malati a unirsi per scambiare esperienze, consigli e idee. Quando si riceve la diagnosi di questa malattia, infatti, si vuole naturalmente vincere l’isolamento e sorge spontaneo il bisogno di condividere il percorso, spesso molto difficile, della diagnosi e, in un secondo momento, quello altrettanto faticoso della terapia.
Obiettivo dell’Associazione è la diffusione della conoscenza della Malattia di Lyme che, nonostante sia classificata come patologia rara, ha visto negli ultimi anni un significativo aumento di casi anche nel nostro Paese, incremento favorito dai cambiamenti climatici e da un sempre più intenso utilizzo del suolo.
È quindi sempre più importante che la cittadinanza conosca come prevenire questa malattia e possa essere supportata una volta eventualmente contratta, ma è ancor più rilevante che il personale sanitario sia in grado di identificare i sintomi di questa patologia difficile da diagnosticare e che, se non riconosciuta tempestivamente, comporta conseguenze spesso gravi e invalidanti sul decorso clinico.
Come Associazione ci impegniamo a:
realizzare campagne di informazione sui media e organizzare convegni, incontri, … rivolti sia alla popolazione sia al personale sanitario, per suscitare una maggiore consapevolezza della Malattia di Lyme e di altre Coinfezioni a essa correlate, ma anche per rendere abituali pratiche di prevenzione efficaci e pervasive;
supportare i malati e le loro famiglie nell’ottenimento di una corretta diagnosi e nel rivolgersi alle professionalità sanitarie più competenti e ai centri di cura all’avanguardia;
sottoscrivere partnership con Enti e Istituzioni di quei Paesi dove la Malattia di Lyme e le altre Coinfezioni sono trattate da più tempo, adottando protocolli di cura ancora non praticati in Italia;
promuovere il riconoscimentosociale e istituzionale della Malattia di Lyme nel nostro Paese, affinché siano garantiti il diritto al lavoro, all’istruzione e all’assistenza psicologica dei malati;
promuovere campagne di fundraising persostenere la ricerca scientifica sulla Malattia di Lyme e sulle patologie correlate;
informare costantemente suiprogressi scientifici raggiunti in Italia e all’estero in merito a questa malattia e divulgare i risultati conseguiti dalla ricerca.
Associazione Lyme Italia e Coinfezioni: si è costituita a Milano con atto notarile nel dicembre 2015 ed è iscritta al Registro Regionale del Volontariato della Lombardia, sezione Provincia Monza e Brianza n. 86/2016 Onlus (Organizzazione non lucrativa di utilità sociale).
Associazione Lyme Italia e Coinfezione ha i seguenti partner istituzionali:
In settembre 2016, durante la “notte Europea dei ricercatori” svoltasi a Torino, (iniziativa promossa e co-finanziata dalla Commissione Europea all’interno del Programma Quadro europeo per la Ricerca e l’Innovazione Horizon 2020) l’associazione ha partecipato come ospite nel “banchetto” dell’Istituto Zooprofilattico del Piemonte, Liguria e Valle d’Aosta, promuovendo in questo modo la conoscenza della stessa e l’informazione della malattia.
In aprile 2017 si è realizzata una campagna di informazione “pensata” per le scuole superiori. Questo progetto è stato realizzato in Piemonte grazie all’importante contributo dell’Istituto Zooprofilattico del Piemonte, Val d’Aosta e Liguria che ha supportato questa azione fornendo il personale formato e qualificato su questa tematica, inserendosi in quella che è l’alternanza scuola-lavoro per le scuole superiori di secondo grado.
Si è realizzata la Brochure prevenzione “Perché fare attenzione alle zecche” con la collaborazione degli Istituti Zooprofillatici del nord Italia, IZTO Istituto Zooprofilattico sperimentale del Piemonte, Liguria e Val D’Aosta, IZSLER Istituto Zooprofilattico sperimentale della Lombardia, Emilia Romagna, IZSVE Istituto Zooprofillatico delle Venezie, Veneto e Friuli Venezia Giulia.
In maggio 2017 Associazione Lyme Italia e coinfezioni ha organizzato con GISML – Gruppo di Studio per la Malattia di Lyme, il V° Congresso Nazionale sulla Malattia di Lyme tenutosi il 27 maggio a Venezia che ha visto l’apprezzato intervento anche del Dr. R. I. Horowitz (U.S.A.) e del professor Luc Montagnier (premio Nobel per la medicina 2008).
In data 26 giugno 2017, L’Associazione ha aderito al GISML – Gruppo di Studio per la Malattia di Lyme.
In data 27 luglio 2017 l’Associazione Lyme ha partecipato alla conferenza di presentazione del III Rapporto MonitoRARE – Camera dei Deputati – Roma Montecitorio. La collaborazione con Alleanza Malattie Rare è proseguita con la partecipazione all’incontro del 9 novembre 2017.
In seguito a una raccolta fondi organizzata nel 2015 con i nostri associati, sono stati acquistati in ottobre 2017 alcuni kit diagnostici, poi donati all’Università di Trieste per condurre uno studio pilota di ricerca di particolari autoanticorpi nel siero di circa 30 soggetti con malattia di Lyme, nei suoi diversi stadi, e con sindrome da malattia di Lyme post-trattamento. Lo scopo della ricerca è quello di identificare nuovi marcatori biologici. È stato già pubblicato – su questo argomento – un articolo sulla rivista scientifica Medical Hypotheses (numero di gennaio 2018), dal titolo: “Autoimmunity against a glycolytic enzyme as a possible cause for persistent symptoms in Lyme disease“.
Grazie alle donazioni dei nostri associati e alla collaborazione della Clinica Dermatologica di Trieste, diretta dal Prof. Giusto Trevisan, si è finanziato un progetto per inviare un medico che lavora presso l’Ospedale Maggiore di Trieste per svolgere un training presso la clinica diretta dal Dr. R.I. Horowitz (USA). Titolo del progetto: ”TREATING RESISTANT LYME AND CHRONIC DISEASE”. Il training si è svolto nel mese di dicembre 2017. Seguirà uno studio su un campione di pazienti affetti da Lyme cronica.
L’associazione si è mossa per diffondere nelle scuole la conoscenza della malattia di Lyme. Il progetto, sebbene poco visibile, coinvolge anche per l’anno scolastico 2017-2018 diverse centinaia di ragazzi degli Istituti d’istruzione superiore di secondo grado della Regione Piemonte. Diffusione di conoscenza, informazione e formazione sono i nodi principali per un’adeguata prevenzione, fattore importantissimo, visti i limiti diagnostici e terapeutici attuali.
Il focus è comunque un insieme più ampio rispetto alle persone affette da malattia di Lyme: giuridicamente l’associazione è un’organizzazione di volontariato, lo stakeholder è tutta la popolazione e non solo il malato.
Monza 1 gennaio 2018
Il Consiglio direttivo di Associazione Lyme Italia e Coinfezioni
I have always loved this speech from the movie Armageddon (see video below), but now I realize that there is an interesting parallelism between this sci-fi movie and what we are living right now with the quest for the solution of ME/CFS and related diseases. Just as the US president says in the movie (where a menacing asteroid is travelling toward the Earth), for the first time in the history of the planet we have the technology to understand what is going on in the bodies and in the brains of ME patients and hopefully the tools to end this tragic loss of lives. And just as it happens in the movie, we have to join our efforts in order to fight this global threat.
A traslation of this blog post to Spanish can be downloaded here. I would like to thank Humbert.Cat for the translation.
1. Introduction
These are some notes about the talk that Mark Davis gave during the Community Symposium held in August at Stanford (video). I will introduce some basic notions about T cell receptors (TCR) in paragraphs 2, 3, 4, and 5. Paragraphs 6 is a description of an innovative technology developed by Mark Davis and his colleagues, based on information gathered from the video itself and three research papers published by Davis and others in the last 4 years. This background should be hopefully enough to allow a good understanding of the exciting pilot data presented by Mark Davis on T cell activity in ME/CFS (paragraph 7), and in chronic Lyme (paragraph 8), and to realize why this technology promises to be some sort of universal test for any kind of infectious and autoimmune diseases, known or unknown.
2. T cells
T cells are a type of leukocytes (also known as white blood cells), the cellular component of our immune system. Most of our circulating T cells are represented by T helper cells (Th cells) and cytotoxic T lymphocytes (CTL). While the function of Th cells is to regulate the activity of other leukocytes through the production of a wide range of chemicals (cytokines), CTLs are directly involved in the killing of host cells infected by pathogens. T cells belong to the adaptive arm of the immune system, along with B cells (the factories of antibodies), and as such, they are meant to provide a defence tailored to specific pathogens: our immune system can provide not only antibodies specific for a given pathogen but also specific T cells (both Th cells and CTLs). The innate arm of the immune system (which includes natural killer cells, macrophages, dendritic cells, mast cells…) on the other hand can provide only a one-fits-all type of defense, which represents the first line of immune response, during an infection.
3. T cell receptor
T cells search for their specific pathogens thanks to a molecule expressed on their surface, called T cell receptor (TCR). In figure 1 you can see a schematic representation of the TCR and of the mechanism by which T cells recognize their targets. Antigens (proteins) from pathogens are presented to T cells by other cells of our body: they are displayed on molecules called major histocompatibility complex (MHC), expressed on the outer membrane; if the antigen fits the TCR of a specific T cell, then this T cell is activated and proliferates (clonal expansion). The two chains (α and β) are assembled using the transcription of gene segments with several copies each: in other words, TCRs are assembled with peptides chosen randomly within a set of several possible choices. This leads to a repertoire of 10^15 possible different TCRs (Mason DA 1998). Each T cell displays only one type of TCR.
Figure 1.Upper half. Th cells and CTLs share the same TCR: in both cases this molecule is the assembly of two peptides (chain α and chain β), but while the TCR of Th cells (on the right) is expressed next to the molecule CD4 (which binds to class II MHC), the TCR of CTL is associated with the molecule CD8 (on the left), which is specific for MHC I. Black bars represent four chains (a complex called CD3) that are involved in the signaling of the TCR with the nucleus of the cell (by Paolo Maccallini). Lower half. A beautiful structural representation of the TCR, bound to the peptide-MHC complex (pMHC), from (Gonzàlez PA et al. 2013). In green the peptide, in blue the β chain, in dark green the α chain. CDRs (complementarity determining regions, orange) are composed of those residues of the α and β chains that directly bind the pMHC.
4. T helper cells
Th cells can recognize only antigens presented by class II MHC: this class of MHC is expressed on the outer membrane of some leukocytes, mainly dendritic cells, B cells, and macrophages (referred to as antigen presenting cells, APCs). MHC II engages the TCR of Th cells thanks to peptide CD4 (expressed exclusively by Th cells). The antigen presented by MHC II is a peptide with a length of 13-17 amino acids (Rudensky, et al., 1991) (figure 2).
Figure 2. The TCR expressed by a Th cell binds an epitope presented by a class II MHC expressed on the plasma membrane of an APC. Chains α and β of MHC II are also represented (by Paolo Maccallini).
5. Cytotoxic T lymphocytes
TCRs expressed by CTLs can bind only antigens displayed by class I MHC, which can be found on the outer membrane of any cell of our body. CD8 is the molecule that makes the TCR expressed by CTLs specific for MHC I. While antigens presented by APCs belongs to pathogens that have been collected on the battlefield of the infection, peptides displayed by class I MHC of a specific cell belong to pathogens that have entered the cell itself, therefore they are the proof of an ongoing intracellular infection (figure 3). When a CTL recognizes an antigen that fits its TCR, then the CTL induces apoptosis (programmed death) of the cell that displays it. Antigens presented by MHC I are peptides in the range of 8 to 10 amino acids (Stern, et al., 1994).
Figure 3. An infected cell displays a viral antigen on its MHC I. The TCR of a CTL binds this peptide and send a signal to the nucleus of the CTL itself, that responds with the induction of apoptosis (releasing granzymes, for instance) of the infected cell (by Paolo Maccallini).
6. The universal immune testing
In his talk, Mark Davis presents an overview of some basic concepts about the immune system, before introducing his exciting new data about ME/CFS and post-treatment Lyme disease syndrome (PTLDS, also known as chronic Lyme). But he also describes a few details of a complex new assay that is theoretically able to read all the information packed in the repertoire of TCRs present – in a given moment – in the blood of a human being. As such, this test – that I have named the universal immune testing – seems to have the potential to determine if a given patient has an ongoing infection (and the exact pathogen) or an autoimmune disease (and the exact autoantigen, i.e. the tissue attached by the immune system). To my understanding, this assay requires three steps, described in the following sections.
6.1. First step: TCR sequencing
As said in paragraph 3, when T cells encounter their specific peptide presented by MHC, they proliferate so that in blood of patients with an ongoing infection (or with a reaction against self, i.e. autoimmunity) we can find several copies of T cells expressing the same TCR: while in healthy controls about 10% of total CD8 T cells is represented by clones of a few different T cells (figure 4, first line), in early Lyme disease – an example of active infection – and in multiple sclerosis (MS) – an example of autoimmune disease – we have a massive clonation of a few lines of CTLs (figure 5, second and third line, respectively). The first step of the universal immune testing is represented by the identification of the exact sequence of TCRs expressed by T cells in blood, as reported in (Han A et al. 2014) where it is described how to sequence genes for the α and the β chain of a given T cell. This approach allows to build graphs of the kind in figure 4, and therefore to determine whether the patient has an abnormal ongoing T cell activity or not. If a clonal expansion is found, then we can speculate that either an active infection is present or some sort of autoimmune condition.
Figure 4. Each circle represents a patient. In the first line, we have four healthy controls, with no clonal expansion of CD8 T cells (the first one, left) or with only a low-level clonal expansion (slices in blue, white, and grey). In the second line, we have four patients with active Lyme disease (early Lyme) and all of them present a massive expansion of only three different T cells (slices in red, blue and orange). In the third line, we have four MS patient with most of their CD8 T cells represented by clones of a bunch of T cells. From the talk by Mark Davis.
6.2. Second step: TCR clustering
Mark Davis and his group have been able to code a software that allows to identify TCRs that share the same antigen, either within an individual or across different donors. This algorithm has been termed GLIPH (grouping of lymphocyte interaction by paratope hotspots) and has been found capable – for instance – to cluster T CD4 cell receptors from 22 subjects with latent M. tuberculosis infection into 16 distinct groups, each of which comprises TCRs from at least 3 different donors (Glanville J et al. 2017). Five of these groups are reported in figure 5. The idea here is that TCRs that belong to the same cluster, react to the same peptide-MHC complex (pMHC).
Figure 5. Five group of TCRs from 22 different donors with latent tuberculosis, clustered by GLIPH. The first column on the left has TCRs IDs, the second one reports donors IDs. Complementarity determining regions (CDR) for the β and the α chains are reported in the third and fifth column, respectively. From (Glanville J et al. 2017).
6.3. Third step: quest for the epitope(s)
As we have seen, this new technology allows to recognize if T cell clonal expansion is an issue in a given patient, by sequencing TCRs from his peripheral blood. It also allows to cluster TCRs either within an individual or across different patients. The next step is to identify what kind of antigen(s) each cluster of TCRs reacts to. In fact, if we could recognize these antigens in a group of patients with similar symptoms, with T cell clonal expansion and similar TCRs, we would be able to understand whether their immune system is fighting a pathogen (and to identify the pathogen) or if it is attacking host tissues and, if that was the case, to identify what tissue. As mentioned, the number of possible TCR gene rearrangement is supposed to be about 10^15, but the number of possible Th cell epitope is about 20^15 which is more than 10^19. This implies that TCRs have to be cross-reactive to some extent, in order to recognize all possible peptides presented by MHCs (Mason DA 1998). The exact extent of this cross-reactivity and the mechanism by which it is obtained has been elucidated by Mark Davis and his colleagues in a recent paper (Birnbaum ME et al. 2014) that gives us the third step of the universal immune testing. The aim of this phase is to take a given TCR and to find the repertoire of his specific antigens (as said, one TCR reacts to several antigens). In order to understand how this is possible let’s consider one of the experiments described in the paper mentioned above. The researchers considered two well-defined TCRs (named Ob.1A12 and Ob.2F3), cloned from a patient with MS and known to recognize peptide 85-99 (figure 6) of myelin basic protein (MBP) presented by HLA-DR15. They then prepared a set of yeast cells expressing HLA-DR15 molecules, each presenting a different peptide of 14 amino acids, with fixed residues only at position 1 and 4, where the peptide is anchored to MHC (figure 6, left). When copies of Ob.1A12 are added to this culture of yeast cells expressing pMHC complexes, they bind only some of them, and as you can see in the right half of figure 6, for each position of the epitopes bound by Ob.1A12, there is an amino acid that is more likely: for instance, the typical epitope of Ob.1A12 preferentially has alanine (A) at position -4, histidine (H) at position -3, arginine (R) at position -2, and so forth. As you can see, histidine (H) at position 2 and phenylalanine (F) at position 3 are obligate amino acids for a Ob.1A12 epitope.
Figure 6. On the left: peptide 85-99 of myelin basic protein (first row) is known to be an epitope for Ob.1A12. At position 1 and 4 it has two residues that allow its binding to the MHC molecule. At position -2, -1, 2, 3, and 5 we find those residues that bind the TCR. The second row represents the generic epitope of the peptide library used to identify the degree of crossreactivity between all the possible Ob.1A12 specific epitopes. On the right: the likelihood of amino acids for each position of Ob.1A12 specific epitopes is represented by shades of violet. As you can see, histidine (H) at position 2 and phenylalanine (F) at position 3 are obligate amino acids for a Ob.1A12 epitope. From (Birnbaum ME et al. 2014).
The table on the right side of figure 6 is, in fact, a substitution matrix with dimension 14×20, a tool that can be used to scan the peptide database in order to find, among all the known peptides expressed by living creatures, all the possible Ob.1A12 specific epitopes. Substitution matrices are commonly used for the so-called peptide alignment, a technique that aims at the identification of similarities between peptides. These matrices are based on evolutionary considerations (Dayhoff, et al., 1978) or on the study of conserved regions in proteins (Henikoff, et al., 1992). But the search for specific epitopes of a given TCR requires (as we have seen here for Ob.1A12) a substitution matrix built ad hoc for that TCR: each TCR requires its own substitution matrix that is obtained adding clones of that TCR on a culture of yeast cells presenting a huge peptide library on their MHCs, and analyzing data from this experiment. So, quite a complex process! In the case of Ob.1A12, this process led to 2330 peptides (including MBP), while the Ob.2F3 specific substitution matrix found 4824 epitopes within the whole peptide database. These peptides included both non-human proteins (bacterial, viral…) and human peptides. For 33 of them (26 non human and 7 human proteins), this group of researchers performed a test in order to directly verify the prediction: 25/26 of environmental peptides and 6/7 of the human peptides induced proliferation of T cells expressing Ob.1A12 and/or Ob.2F3, and this is a huge proof of the validity of this analysis! These 33 peptides are reported in figure 7. This is the last step of the universal immune testing, the one that from the TCR leads to the epitopes. As you have seen, a huge set of different peptides from different sources is linked to each single TCR, in other words, crossreactivity is an intrinsic property of TCR. This also means that lymphocyte transformation tests (LTTs), widely used in Europe for the detection of infections like Borrelia burgdorferi and others, bear a high risk of false-positive results and require a process of experimental and theoretical validation, that is currently lacking (see also this post on this issue).
Figure 7. A set of 33 peptides (both human and environmental) predicted to be specific epitopes for both Ob.1A12 and Ob.2F3. From (Birnbaum ME et al. 2014).
We are now ready to fully appreciate the pilot data that Mark Davis presented at the Symposium on CD8 T cell clonal expansion in ME/CFS and in chronic Lyme.
7. We have a hit!
Mark Davis, along with Jacob Glanville and José Montoya, has sequenced TCRs from the peripheral blood of 50 ME/CFS patients and 49 controls (first step of the universal immune testing, remember?), then they have clustered them using the GLIPH algorithm (second step). They have found 28 clusters with more than 2500 similar sequences each, where each cluster collects multiple sequences from the same individual as well as (which is perhaps more important) sequences from different patients (figure 8). The cluster that I have circled in red, for instance, is a collection of more than 3000 similar TCRs. The presence of this wide clusters in ME/CFS patients, compared to healthy controls, represents an indirect proof of a specific T cell response to some common trigger in this group of patients, which might be a pathogen or a tissue of the body (or both).
Figure 8. In ME/CFS, TCRs sequences from 50 patients form 28 clusters with more than 2500 similar sequences, and this is clearly not the case in healthy controls. This point to some specific immune response to a pathogen or to a human tissue (or both). This slide is from the talk by Mark Davis.
Among these 50 ME/CFS patients, Davis and colleagues selected 6 patients with similar HLA genes (figure 9, left), 5 females among them, and studied their TCRs deeper. In the right half of figure 9, you can see for each patient the degree of CTL clonal expansion. Remember that in healthy controls only about 10% of CTLs is composed by clones of a few cells (figure 4, first raw), while here we see that about 50% of all CTLs is composed by clones. So, a “marked clonal expansion” of CD8 T cells, as Davis said.
Figure 9. On the left: 6 MECFS patients with similar HLA genes have been selected. Patient ID is reported in the first column on the left, the second column indicates the age of each patient, the third indicates the gender, the fourth column is about exposure to cytomegalovirus, the third one is on MHC I genes. On the right: analysis of clonal expansion of CD8 T cells for each of the six patients. There is a marked clonal expansion (about 50%) compared to healthy controls (about 10%).
Sequences of α and β chains of TCRs from three of the six patients (patients L4-02, L4-10, and L3-20) are reported in figure 10 (I have verified that in fact these are sequences of α and β chains of human TCRs using them as query sequences in standard protein BLAST).
Figure 10. Beta chains (first column) and respective α chains (fifth column) from 3 ME/CFS patients (L4-02, L4-10, and L3-20, last column).
So, we have seen so far the first two steps of the universal immune testing in ME. What about the third step? In his talk, Mark Davis didn’t present any particular epitope, he just showed a slide with what likely is the selection of the epitopes from the peptide library (see paragraph 6.3) by one of the TCRs reported in figure 10. This selection is reported in figure 11, but from that picture, it is not possible to gather any information about the identity of these epitopes. As you probably remember from paragraph 6.3, the analysis of the peptides selected by a TCR among the peptide library allows the identification of a substitution matrix that can be used to select all the possible epitopes of that specific TCR, from the peptide database. This last crucial step has to be performed yet, or it has been already performed, but Davis has not communicated the preliminary results during his talk. Recently new resources have been made available by Open Medicine Foundation, for this promising research to be further pursued, among other projects (R). The aim here, as already said, is to find the antigen that triggers this T cell response. As Mark Davis said, it might be an antigen from a specific pathogen (perhaps a common pathogen that comes and goes) that elicits an abnormal immune response which ends targeting some host tissue (microglia, for instance), thus leading to the kind of immune activation that has been recently reported by Mark Davis himself and others in ME/CFS (Montoya JG et al. 2017). The idea of a common pathogen triggering a pathologic immune response is not new in medicine, and rheumatic fever (RF) is an example of such a disease: RF is an autoimmune disease that attacks heart, brain and joints and is generally triggered by a streptococcal throat infection (Marijon E et al. 2012). The other possible avenue is, of course, that of an ongoing infection of some kind, that has yet to be detected. As said (see par. 6.1), CD8 T cell clonal expansion is present in both acute infections (like early Lyme disease) and autoimmune diseases (like MS) (figure 4), so we have to wait for the antigen identification if we want to understand if the CTLs activity is against a pathogen and/or against a host tissue.
Figure 11. In this picture, we can see the selection, through several rounds, of a bunch of peptides by a particular TCR from a ME patient. The selection takes place among a huge collection of peptides presented by HLA-A2 (MHC I) expressed by yeast cells. At each round the number of possible peptides is smaller.
8. Chronic Lyme does exist
It has probably been overlooked that in his talk, Mark Davis reported also very interesting data on post-treatment Lyme disease syndrome (PTLDS, also known as chronic Lyme disease). In particular, he found a marked clonal expansion in CD8 T cells of 4 PTLDS patients (about 40% of total CTLs) as reported in figure 12: consider that in this case, blue slices represent unique T cells, while all the other slices represent clones! All that has been said about CD8 clonal expansion in ME/CFS does apply in this case too: it might be the proof of an ongoing infection – perhaps the same B. burgdorferi, as suggested by several animal models (Embers ME et al. 2017), (Embers ME et al. 2012), (Hodzic E et al. 2008), (Yrjänäinen H et al. 2010) – or a coinfection (a virus?) or it could be the expression of an autoimmune reaction triggered by the initial infection. This has still to be discovered, running the complete universal immune testing, but what is already clear from figure 12 is that PTLDS is a real condition, with something really wrong going on within the immune response: chronic Lyme does exist.
Figure 12. CD8 T cells clonal expansion in four chronic Lyme patients: there is a marked clonal expansion that stands for T cell activity against a pathogen or against host tissue.
9. Conclusions
Mark Davis and other researchers have developed a complex assay that is able to sequence TCRs from patients, cluster them into groups of TCRs that react to the same antigens, and discover the antigens that triggered that particular T cell response. This assay is a kind of universal immune testing that is theoretically able to recognize if a person (or a group of patients) presents an immune response against a pathogen or against one of his own tissues (or both). This approach has already given pilot data on an ongoing CD8 T cell activity in ME/CFS patients and in chronic Lyme patients and will hopefully identify the trigger of this immune response in the near future. Whether ME/CFS is an ongoing infection, an autoimmune disease or both, the universal immune testing might be able to tell us. This new technology is for immunology, what whole genome sequencing is for genetics, or metabolomics is for molecular diseases: it doesn’t search for a particular pathogen or a particular autoimmune disease. No, it searches for all possible infections and immune disorders, even those that have yet to be discovered.
Monica Embers (Tulane University) pubblica due nuovi studi sul modello animale di Lyme tardiva (R), (R).
Dieci macachi (Macaca mulatta) sono stati infettati con B. burgdorferi. Dopo 5 mesi (Lyme tardiva) sono stati trattati con doxyciclina orale (5 mg/kg) per un mese. Dopo altri 8 mesi sono stati sacrificati e diversi campioni di tessuto sono stati sottoposti a svariate indagini che hanno dimostrato la presenza di fenomeni infiammatori (infiltrati linfocitari) in corrispondenza di rare spirochete (figura 1). Questo macaco condivide con gli esseri umani il 97.5% del DNA (R) ed è dunque presumibilmente un buon modello animale di malattia di Lyme.
Figura 1. Presenza di spirochete residue negli animali trattati, localizzate nel neuropilo cerebrale (regione compresa tra i corpi cellulari dei neuroni) (F), nel tronco encefalico (G) e in un nervo periferico della gamba (K).
Lo studio dimostra anche la esistenza di una infezione residua in due esemplari sieronegativi, confermando la nozione secondo la quale la sierologia può essere poco sensibile non solo nella fase iniziale (cosa ben nota e riconosciuta), ma anche nella fase tardiva della infezione (figura2). Si evince anche che la risposta immunitaria al C6, a volte usata come test di primo livello, non rimane positiva in tutti i casi di infezione persistente.
Figura 2. Sopra la risposta anticorpale alle principali proteine immunogeniche di B.b. in 4 dei 10 individui studiati. Sotto il risultato di alcune indagini negli stessi individui. In rosso un individuo che risulta positivo al test della xenodiagnosi, ma è tuttavia sieronegativo. In verde un individuo che perde la sua risposta immunitaria nel tempo, pur essendo positivo alla xenodiagnosi. In blu un individuo che perde la risposta al C6, pur essendo positivo a diversi test diretti.
Secondo gli autori, questi dati suggeriscono che i sintomi cronici della malattia di Lyme siano dovuti alla sopravvivenza di alcuni esemplari di Borrelia in vari tessuti. Lo studio conferma un lavoro precedente dello stesso gruppo, sullo stesso modello animale (Embers, 2012). Risultati analoghi sono stati riportati in altri modelli animali (Hodzic, E et al. 2008), (Yrjänäinen, 2010).
Mi è stato chiesto di scrivere due righe che potessero sintetizzare quali sono i problemi attuali nella malattia di Lyme e quali dovrebbero essere i campi di intervento per migliorare la situazione. Questi argomenti sono trattati nel blog, con un ampio corredo bibliografico. Ma volendo sintetizzare, ciò che segue è quello che penso.
“Il problema maggiore nella malattia di Lyme attualmente è quello rappresentato dai pazienti con sintomi cronici. Per questo gruppo di pazienti (che ricade sotto la entità nosografica denominata post-treatment Lyme disease syndrome, PTLDS) non esistono terapie approvate, in nessuna nazione europea e – a quanto ne so – nel mondo.
I pazienti sono da un lato sminuiti nella loro sofferenza da medici che negano l’obiettività dei sintomi cronici e dall’altro sono oggetto di vere e proprie truffe da parte di cliniche private che offrono test privi di validazione scientifica e terapie di efficacia non dimostrata e potenzialmente dannose.
Non è nota la causa della persistenza dei sintomi dopo le terapie raccomandate e le ipotesi considerate sono: persistenza della infezione, presenza di altri agenti patogeni non identificati e non trattati, disfunzioni immunitarie, danni residui ai tessuti, danno al microbiota. Nessuna di queste ipotesi è stata dimostrata in modo convincente.
Penso sia un dovere occuparci di questi pazienti, fino ad oggi completamente trascurati dalla medicina e rifiutati dal sistema sanitario. Sarebbe auspicabile una strategia di ricerca aggressiva che investighi i seguenti punti:
lo sviluppo di un test per la infezione attiva da Borrelia burgdorferi;
la ricerca della eziologia dei sintomi cronici, attraverso indagini immunologiche, metabolomiche, metagenomiche, genomiche, proteomiche, di espressione genica, di brain imaging etc;
trial clinici con terapie innovative per la fase cronica;
studi di microbiologia sul fenomeno della persistenza batterica, ormai ampiamente documentato in vitro nel caso di Borrelia burgdorferi (R);
studi di neuroimmunologia che approfondiscano il dato riportato in due ricerche di ampio respiro, secondo il quale il 50% di questi pazienti presentano anticorpi contro il sistema nervoso centrale e periferico (R).
Studi su questi ambiti sono già in corso negli USA, ma date le specificità europee dell’agente patogeno e delle manifestazioni cliniche, è possibile che i risultati ottenuti nel Nuovo Mondo non siano del tutto applicabili ai pazienti europei ed è pertanto necessario che le nostre università conducano le proprie investigazioni. Non ci si può adagiare sui risultati prodotti oltreoceano.
Bisogna incoraggiare i giovani ricercatori a scegliere questo campo per la loro carriera, presentando la malattia cronica come un problema insoluto e difficile da risolvere, ma affascinante in quanto interdisciplinare, a cavallo tra la microbiologia, l’immunologia, le neuroscienze, lo studio del metabolismo, la ingegneria degli strumenti diagnostici, la bioinformatica etc. Si deve inoltre sottolineare che lo studio di questa condizione cronica potrebbe avere ricadute positive su altre malattie, come ad esempio quelle neuroimmuni, con prospettive di grandi benefici per la collettività, che possono esulare dall’ambito della PTLDS.”